Biologie du neurone - Electrophysiologie
2.6.- LA BIOLOGIE DU NEURONE
2.6.1.- METABOLISME DU NEURONE
Le métabolisme du neurone est semblable à celui des autres cellules. Mais étant très différencié il n’est pas capable de multiplication.
Son métabolisme est principalement basé sur la présence de glucose, d’oxygène et de co-facteurs vitaminiques. Deux substances lui sont spécialement indispensables : ce sont les vitamines B1 (ou Thiamine) et PP (Pellagra Preventing). La vitamine B1 assure l’utilisation complète des glucides et son absence provoque des neuropathies carentielles avec sclérose et dégénérescence des neurones. L’avitaminose PP, appelée Pellagre, provoque chez l’homme des troubles nerveux et mentaux par neuropathie et encéphalopathies carentielles.
Le neurone respire activement et il est très sensible à la privation d’oxygène. Les neurones du cerveau sont les plus vulnérables à l’anoxie et 3 minutes d’anoxie totale peuvent provoquer des lésions cérébrales irréversibles..
2.6.2. - LA DEGENERESCENCE WALLERIENNE
Lorsque le corps cellulaire d’un neurone est détruit, la fibre nerveuse dégénère totalement. Lorsque la fibre nerveuse est sectionnée, la partie distale, c’est à dire la partie qui est séparée du corps cellulaire, dégénère. C’est le phénomène de la dégénérescence wallérienne. Par contre la partie proximale, c’est à dire celle qui est restée en rapport avec le corps cellulaire, peut régénérer
2.6.3. - LA REGENERATION NERVEUSE S.07
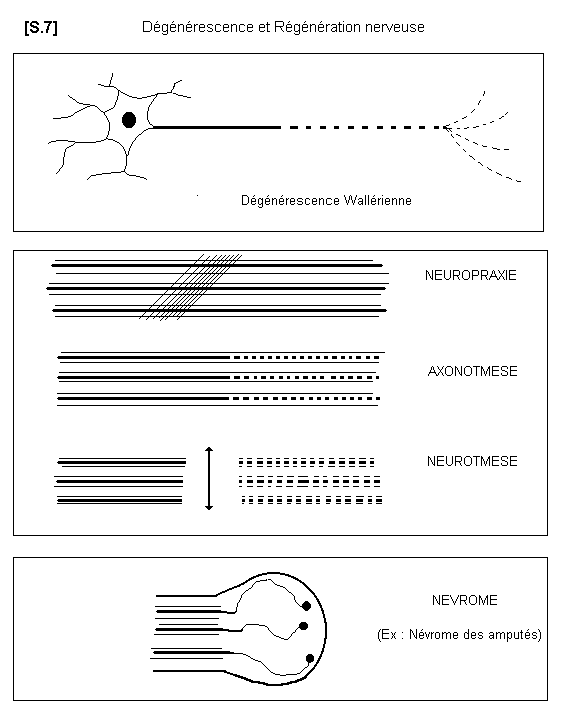
La régénération nerveuse peut se présenter de différentes façons. Si la fibre nerveuse n’est pas sectionnée, mais comprimée (neuropraxie par hématome ou oedème interstitiel), l’influx nerveux est temporairement interrompu. La récupération spontanée survient dans un délai de 6 à 8 semaines.
Si la fibre nerveuse est interrompue avec conservation de ses gaines de SCHWANN et de myéline, la régénération se fait normalement à la vitesse approximative d’un millimètre par jour, et la fibre retrouvera son trajet normal en suivant les gaines.
Si les gaines sont interrompues, la fibre nerveuse peut repousser dans des gaines de voisinage et présenter éventuellement des fausses routes.
Si les gaines sont interrompues et les deux segments très écartés, les fibres n’étant plus guidées se recourbent en pelotons ou en boules appelés névromes, qui sont spécialement douloureux.
On distingue donc ainsi trois grands types de lésions nerveuses périphériques : ce sont la neuropraxie, l’axnotmésis, et le neurotmésis.
2.6.4.- ARTICULATIONS ENTRE LES NEURONES : LES SYNAPSES S.08
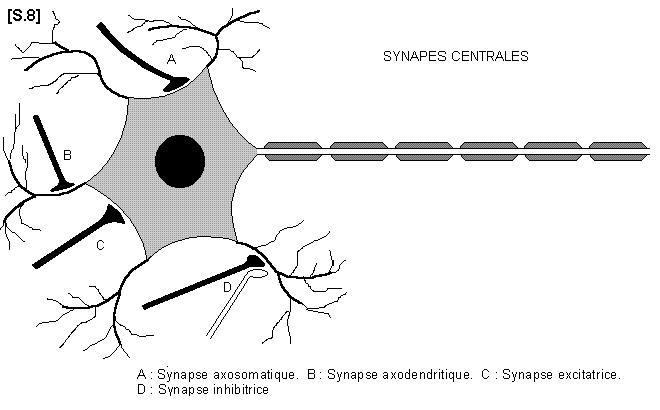
a)- Synapses centrales :
L’ensemble du système nerveux central est constitué de multiples chaînes de neurones disposées bout à bout et en relation réciproque par leur pôle contraire. L’arborisation de l’axone est en effet en rapport avec les dendrites ou le corps du neurone suivant. Les connexions entre les neurones sont appelées synapses. Il existe donc des synapses axo-somatiques et des synapses axo-dendritiques. Certaines sont excitatrices, et d’autres, inhibitrices, assurant ainsi la régulation de la transmission synaptique.
Notion de densité synaptique : il existe environ 6000 synapses pour un motoneurone.
La transmission de l’influx nerveux dans la synapse se fait grâce à la libération locale de substance chimique (adrénaline, acétylcholine, dopamine etc...). Ces substances sont appelées médiateurs chimiques ou neuro-transmetteurs.
b)- Synapses périphériques : Elles sont représentées par la synapse neuro - musculaire de la plaque motrice des fibres musculaires.
2.7.-PROPRIETES ELECTROPHYSIOLOGIQUES DU NEURONE :
INFLUX NERVEUX
2.7.1.- Mise en évidence de l’influx nerveux
L’influx nerveux résulte d’une variation transitoire, de l’ordre d’une milliseconde, de la répartion des ions situés de part et d’autre de la membrane cellulaire.
Dans le cas d’un neurone sensitif, l’influx nerveux prend naissance au niveau d’un récepteur périphérique et se propage le long de la fibre nerveuse jusqu’à son arborisation terminale. La naissance de l’influx nerveux est donc la conséquence de phénomènes physico-chimiques qui ont lieu au niveau du récepteur.
Dans le cas d’un neurone moteur, l’influx nerveux prend naissance à la jonction du corps cellulaire et de l’axone. Il est la conséquence de phénomènes physico-chimiques intervenus au niveau du corps cellulaire à la suite d’une stimulation du neurone moteur par un autre neurone.
On peut mettre en évidence l’existence de l’influx nerveux de la façon suivante :
Chez un animal dont on a dénudé un nerf moteur, les contractions musculaires, volontaires, spontanées, ou stimulées, persistent tout le temps que le nerf n’a pas été écrasé, ligaturé ou sectionné. Les nerfs sont donc parcourus par "quelque chose" que GALIEN au IIème siècle avant Jésus-Christ, appelait "un fluide". Bien que les nerfs et les muscles soient très sensibles au courant électrique et bien que l’on puisse enregistrer à leur contact des phénomènes électriques, l’influx nerveux n’est pas un courant électrique. Ceci est démontré par les deux faits suivants.
a) - un nerf écrasé qui ne conduit plus l’influx nerveux transmet encore, par diffusion, des courants électriques qui sont enregistrables sur le nerf en aval de l’écrasement mais sans effet moteur sur le muscle.
b) - un nerf intact conduit le courant électrique à la vitesse de la lumière (300.000 kilomètres/seconde) alors que l’influx nerveux n’est transmis qu’à une vitesse comprise entre 1 et 100 m/seconde (en fait chez l’homme 49m/s pour les nerfs du membre supérieur et 42m/s pour les nerfs du membre inférieur).
Quoique l’influx nerveux ne soit pas un courant électrique on peut enregistrer le passage de l’influx, car son existence est liée à la production d’un champ électrique. Celui-ci est de faible intensité et l’enregistrement nécessite des amplificateurs et des galvanomètres très sensibles (oscilloscope à tube cathodique).
On peut enregistrer et étudier ainsi soit un potentiel d’action émis au cours du fonctionnement normal du système nerveux, soit un potentiel d’action provoqué par une stimulation électrique portée sur le nerf.
Sur l’oscilloscope cathodique on obtient un premier accident de la ligne de base isoélectrique que l’on appelle "artéfact de stimulation" qui est pratiquement contemporain de la stimulation. On obtient ensuite un second accident appelé potentiel d’action qui répond au passage de l’influx nerveux. Il apparaît avec un retard de quelques millisecondes. En mesurant le temps qui sépare l’artéfact et le potentiel d’action, en connaissant la longueur parcourue sur le nerf, on peut mesurer la vitesse de la conduction nerveuse (motrice ou sensitive). Toutes ces explorations se font dans les services de Neurophysiologie clinique ( Explorations fonctionnelles de Système nerveux).
2.7.2.- Polarisation de la fibre nerveuse au repos : potentiel de membrane ou
potentiel de repos. S.09
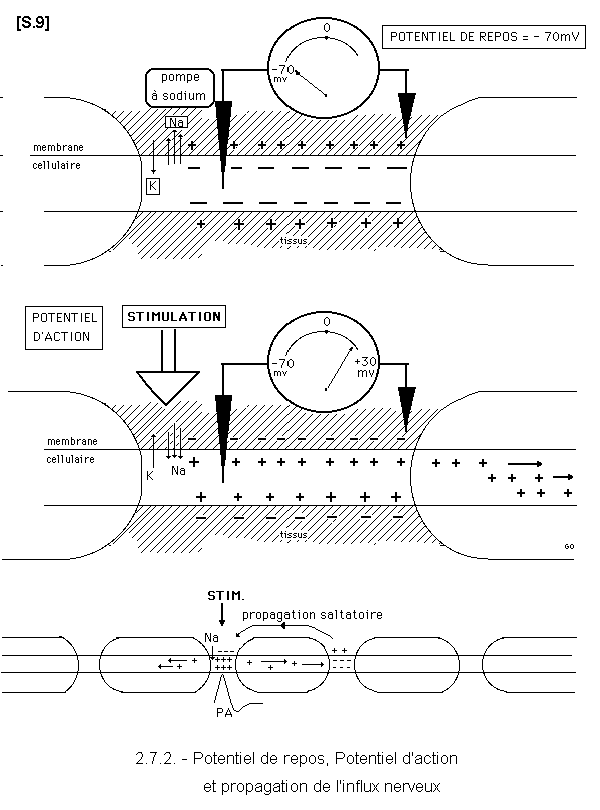
Comme toute structure vivante, la fibre nerveuse est polarisée. Si on introduit à l’intérieur de la fibre une micro-électrode on enregistre, entre celle-ci et une électrode extérieure, une différence de potentiel. Cette différence apparaît brusquement au moment où, ayant franchi la membrane, la micro-électrode pénètre à l’intérieur de la fibre.
L’intérieur de la fibre est négatif par rapport à sa surface. La différence de potentiel est de -70 à -80 millivolts environ. C’est le potentiel de membrane ou potentiel de repos. Le potentiel de repos enregistrable sur toute cellule vivante, s’explique par la théorie ionique de l’influx nerveux de HODGKIN et résulte d’une inégale répartition des ions de part et d’autre de la membrane cellulaire.
Elle est la conséquence de deux sortes de phénomènes :
a) - les uns sont passifs et ne consomment pas d’énergie. Ils correspondent aux lois simples de l’osmose s’appliquant aux membranes semi-perméables.
b) - les autres sont des transports ioniques actifs et consomment de l’énergie qui est fournie par le métabolisme du glucose. Leur rôle est de faire entrer quelques ions K dans la cellule et de chasser de nombreux ions Na hors de la cellule. Ce mécanisme actif du rejet du sodium, qui est permanent au repos, est appelé POMPE A SODIUM.
De ces mouvements permanents il résulte une stabilité, un équilibre de concentration ionique à l’intérieur et à l’extérieur de la fibre, la plus grande concentration d’ions Na se faisant hors de la cellule. Ainsi, au repos, l’extérieur de la cellule est positif et l’intérieur de la cellule est négatif.
2.7.3. Dépolarisation de la fibre nerveuse active : potentiel d’action S.10
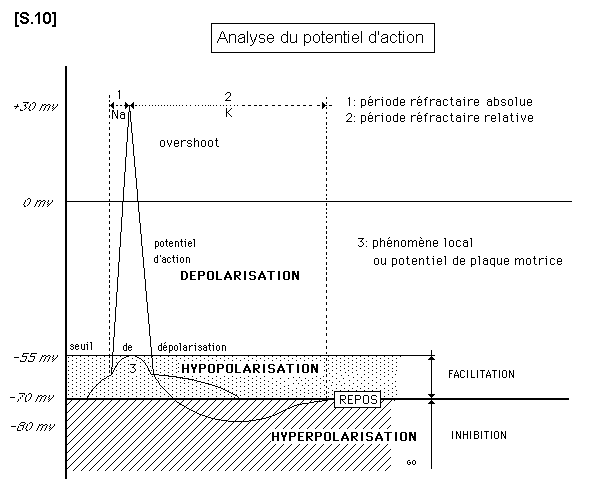
Lorsque la membrane est soumise à une stimulation électrique ou à l’action d’un neurotransmetteur chimique, le rejet actif du sodium diminue, cesse, puis s’inverse. La concentration de Na intra-cellulaire augmente (blocage de la pompe à sodium). D’autre part, la vitesse de passage des ions K à travers la membrane cellulaire est plus petite que la vitesse de passage des ions Na. En conséquence une certaine quantité d’ions Na pénètre dans la cellule avant qu’une égale quantité d’ions K n’en sorte.
L’équilibre est temporairement rompu. On assiste à une dépolarisation de la fibre nerveuse qui se traduit par l’apparition d’un potentiel d’actions (voir schéma). Lorsque le potentiel de membrane a retrouvé son niveau de repos, l’activité de la pompe à sodium reprend. C’est un temps consommateur d’énergie. On dit que la cellule "recharge ses batteries".
Etude des circonstances d’apparition du potentiel d’action
Lorsque la stimulation est faible, ou au début de la stimulation on assiste à une légère diminution de la différence de potentiel. Ceci se traduit sur l’écran par une inflexion appelée "phénomène local". Lorsque l’intensité de la stimulation augmente, l’amplitude du phénomène local atteint le seuil de dépolarisation ( -55 millivolts). Alors éclate un potentiel d’action qui atteint O millivolt et le plus souvent les dépasse. Cette dépolarisation brutale est responsable de l’apparition du potentiel d’action qui est complet, constant, de même amplitude, de même forme et de même durée pour une fibre donnée : c’est la "loi du tout ou rien". La partie ascendante correspond à la pénétration rapide des ions Na, la partie descendante plus lente, correspond à la sortie des ions K. Le potentiel d’action va se propager le long de la fibre nerveuse et sa vitesse de propagation est constante.
2.7.4.- La propagation de l’influx
2.7.4.1.- le long de la fibre nerveuse
La gaine de myéline constitue un isolant mais elle présente de distance en distance des interruptions complètes, ce sont les étranglements de RANVIER. A ce niveau, l’axone est seulement recouvert par la gaine de SWANN qui est perméable aux échanges ioniques. C’est seulement au niveau des étranglements de RANVIER que peuvent s’effectuer les échanges ioniques du potentiel d’action. Au point de stimulation, l’intérieur de la fibre nerveuse est très positif et les ions positifs se répandent de part et d’autre du site de stimulation vers les autres étranglements de RANVIER. Leur présence provoque l’ouverture des pores de la membrane cellulaire permettant à nouveau la pénétration d’ions sodium et une dépolarisation qui provoque un autre potentiel d’action. Ainsi, de segment en segment, le potentiel d’action est entretenu tout le long de la fibre nerveuse. L’influx nerveux parcourt toute la fibre nerveuse par propagation saltatoire, et atteint l’arborisation terminale. La transmission de l’influx se fait à vitesse constante le long d’une fibre nerveuse normale.
2.7.4.2.- au niveau des synapses
Au niveau du franchissement des synapses, cette transmission peut subir trois types de modification appelés facilitation, inhibition, occlusion.
a) - Facilitation : un neurone qui est stimulé et qui va répondre par un potentiel d’action , produit dans son environnement un champ électrique décroissant. Des neurones de voisinage sont soumis à ce champ électrique infra-liminaire, insuffisant pour provoquer la stimulation des neurones voisins. Cependant, si dans le même temps, un autre neurone est lui-même directement stimulé, son champ électrique décroissant va s’étendre aux neurones périphériques qui, bien que n’étant pas stimulés directement, atteignent, de ce fait, le seuil de dépolarisation et sont alors l’objet d’une dépolarisation complète responsable de leur potentiel d’action. On voit que schématiquement deux neurones peuvent être stimulés directement et que plusieurs autres neurones peuvent répondre, par diffusion de l’influx.
b) - Inhibition : certaines articulations synaptiques libèrent des médiateurs chimiques qui augmentent la fuite des ions K alors que la membrane cellulaire reste imperméable au sodium. Il existe de ce fait une hyperpolarisation ( -80 millivolts). Il faut dans ce cas un stimulus plus intense pour faire apparaître une dépolarisation et un influx au niveau des neurones concernés.
c) - Occlusion : lorsque plusieurs neurones reçoivent une double stimulation synchrone, leur fonctionnement est bloqué et il n’y a pas à leur niveau de formation de potentiel d’action. Dans ce cas, deux neurones synchrones peuvent bloquer le fonctionnement de plusieurs autres neurones.
Pr Gérard Outrequin - Dr Bertrand Boutillier - Toute reproduction interdite
Licence d'utilisation - Mentions légales - Politique de confidentialité - Faire un don - Découvrez MedShake !