Le Cerveau (2)
4.4.- Systématisation du cerveau
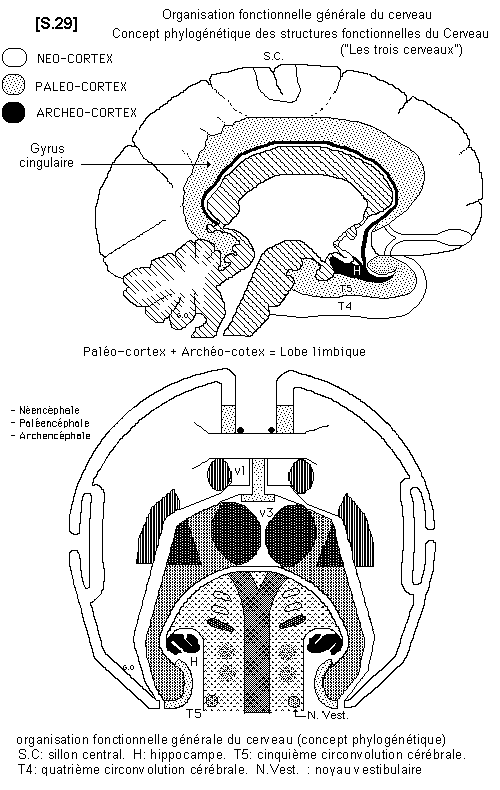
4.4.1.- Organisation fonctionnelle générale (Phylogenèse)
Les centres nerveux cérébraux sont hiérarchisés. Ainsi les centres sous-corticaux sont sous la dépendance du cortex. Cependant les centres sous-corticaux sont capables d’autonomie relative lorsqu’ils assurent des fonctions automatiques. Cette subordination est la conséquence des acquisitions fonctionnelles successives des étapes de la phylogenèse (évolution organique et fonctionnelle, étudiée tout au long des espèces animales successives).
Les structures les plus récentes dominent les structures qui les ont précédées mais celles-ci conservent leurs fonctions propres.
Sur le plan fonctionnel, il est donc intéressant de retrouver, dans les structures du cerveau, les trois niveaux de développement évolutif du Système Nerveux Central (archencéphale : cerveau instinctif et réflexe, paléencéphale : cerveau impulsif et automatique, néencéphale : cerveau conscient puis rationnel, chez l’homme). A chaque stade évolutif, le cerveau possède un cortex et des noyaux gris.
4.4.1.1.- Archencéphale : Cerveau instinctif et réflexe
C’est le niveau de développement des poissons et des amphibiens. Les informations sont reçues par le cerveau sensitif et sensoriel (olfactif et visuel). Les centres d’intégration sont représentés par l’archéo-cortex (dont les structures sont présentes chez l’homme : hippocampe dorsal - atrophique chez l’homme - et hippocampe ventral) et la substance réticulée. Le cerveau moteur est représenté par les noyaux sous-opto-striés et les noyaux du toit du mésencéphale (qui correspondent chez l’homme aux tubercules quadrijumeaux). Les voies vestibulaires appartiennent aussi à ce stade de développement.
Les voies motrices peuvent être regroupées sous le nom de Système archéo-moteur qui assure la mobilité ainsi que le positionnement propre à l’espèce (équilibration).
Ainsi à l’origine même du comportement moteur, se trouvent des stimulations sensorielles et des réponses instinctives de nature purement réflexe.
4.4.1.2. - le Paléencéphale : Cerveau impulsif et automatique
C’est le stade de développement des reptiles puis des oiseaux. Il se superpose aux structures précédentes pour constituer le RHINENCEPHALE.
Il comprend un cortex (paléo-cortex), représenté par le gyrus cingulaire et la circonvolution entorhinale ou T5. L’ensemble a la forme classique d’une raquette, dont le manche est constitué par l’appareil olfactif. Il contient, en plus, un centre sensitif et sensoriel, le thalamus, (noyau ventro- latéral postérieur). Les centres moteurs sont constitués par le paléo-striatum (pallidum et noyaux moteurs du thalamus). Ces centres sont reliés entre eux par les voies thalamo-striées. Les voies motrices paléo-striées se projettent sur les noyaux sous-opto-striés. Les voies motrices, à ce niveau de développement, peuvent être regroupées sous le nom de Système paléo-moteur.
Globalement, le Paléencéphale est un cerveau fonctionnellement très complet. Dépassant les impératifs biologiques élémentaires de l’instinct, il assure toute l’activité automatique (mouvement et posture) du comportement.
4.4.1.3.- le Néencéphale : Cerveau conscient puis rationnel chez l’homme
C’est le stade évolutif des mammifères avec des graduations à l’intérieur du groupe. Il est spécialement développé, en volume et en fonction, chez les primates et surtout chez l’homme. Il se superpose aux deux stades précédents, dont les structures restent cependant, fonctionnelles chez l’homme. Il comprend le cortex cérébral. Au plan moteur, ce néo-cortex ou néo-pallium possède lui-même des aires associatives motrices dont les voies peuvent être regroupées sous le nom de Système néo-moteur.
Il possède aussi des aires primaires (aire 4) dont la voie efférente constitue le faisceau pyramidal. Le néencéphale possède ses propres noyaux gris, qui sont le noyau caudé et le putamen (néo-striatum). Par des voies cortico-striées, il contrôle le paléencéphale.
En bref, le concept fonctionnel du cerveau, basé sur la phylogenèse, reconnaît trois structures superposées :
– le cerveau instinctif et réflexe (archencéphale)
– le cerveau impulsif et automatique (paléencéphale)
– le cerveau conscient, puis rationnel (néencéphale)
On retrouve, à tous les niveaux, le comportement fondamental de la matière vivante : Information - Traitement - Action
4.4.2. - Organisation fonctionnelle particulière : Topographie fonctionnelle du cortex
Il existe au niveau de la surface du cortex une disposition topographique des fonctions motrices, sensitives, sensorielles et associatives. Cette organisation des grandes fonctions a été récemment remise en cause par les images obtenues en IRM fonctionnelle, mais les bases de la topographie corticale sont confirmées par la physio-pathologie neurologique et par la clinique.
« La carte cyto-architectonique du cortex cérébral, proposée par Brodmann en 1909, reste le découpage du cortex cérébral le plus utilisé » (Source : Pritchard/Halloway. Neurosciences médicales. 2002. P.229). C’est cette description, toujours classique, qui est exposée ici.
4.4.2.1. Systématisation du néencéphale (Topographie corticale)
4.4.2.1.1. - Fonctions motrices et territoires moteurs S.30
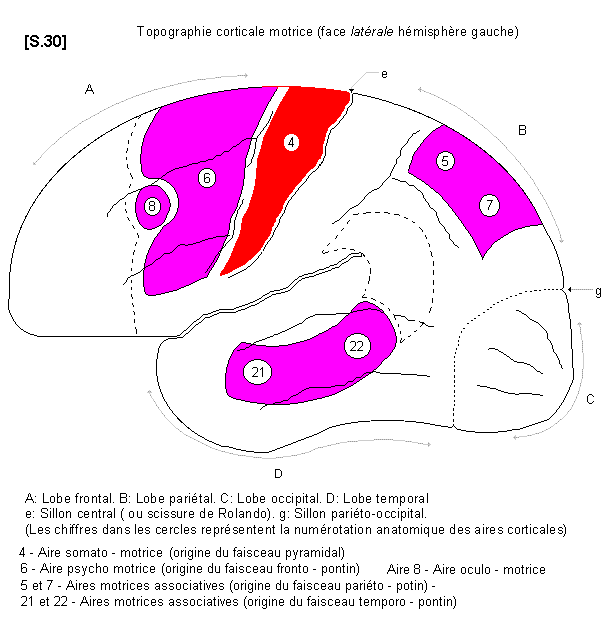
a-Aire 4 (aire somato - motrice:Topographie corticale
L’aire 4 correspond à la plus grande partie du gyrus pré-central (circonvolution frontale ascendante). A son niveau sont situées les grandes cellules pyramidales de BETZ qui sont le point de départ des axones qui constituent le faisceau pyramidal. A noter que la partie la plus élevée de l’aire 4 se prolonge sur la face médiale de l’hémisphère. Tout au long de l’aire 4 il existe une véritable somatotopie fonctionnelle. Chacune des régions de l’aire 4 représente un territoire corporel dont la surface corticale est d’autant plus grande que ce territoire a une grande importance fonctionnelle. Ainsi la surface correspondant à la main est très grande et, plus spécialement, la surface correspondant au pouce. Cette représentation schématique du corps humain à la surface du cortex moteur s’appelle "homonculus de Penfield" S.31
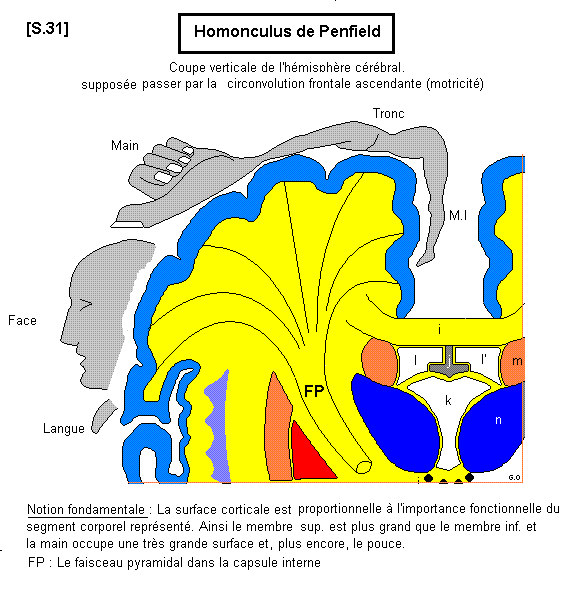
La structure histologique de l’aire 4 est particulière. Elle ne contient pas de cellules sensitives, mais seulement des cellules motrices pyramidales, grandes et petites, et les couches 3 et 5 envahissent toute l’épaisseur du cortex (3mm). Seules les fibres motrices cortico- spinales directes naissent des grandes cellules pyramidales de Betz (100 µ). Elles sont peu nombreuses (35000).Leur axone représente seulement 2 à 3% du faisceau pyramidal. Elles assurent les fonctions motrices distales de précision.
Les autres fibres motrices sont les axones des petites cellules pyramidales dont 60% proviennent de l’aire 4 (faisceau para - pyramidal ou voie adversive). Les 40% restants proviennent des aires motrices associatives du cortex. Globalement, le faisceau pyramidal compte, au niveau des pyramides de la moelle allongée, un million de fibres. Ainsi constituée, la voie pyramidale est donc hétérogène.
* Aire motrice supplémentaire S.32
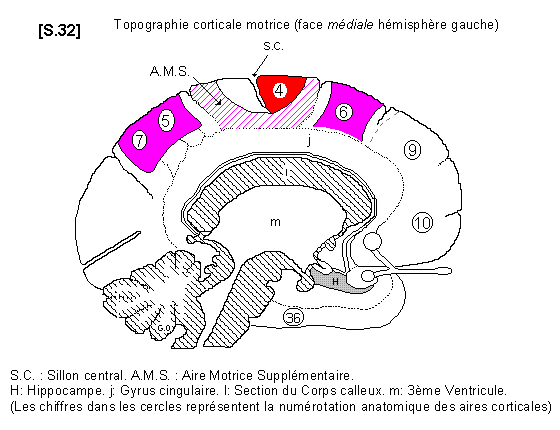
Au niveau du lobule para-central (face médiale de l’hémisphère), il existe un territoire moteur particulier appelé aire motrice supplémentaire (AMS). Elle possède, comme l’aire 4, une somatotopie complète. C’est une aire motrice associative qui parait se rapporter au projet du mouvement (prévision et représentation du mouvement à exécuter).
L’EEG a montré qu’il existe une activité corticale précédant le mouvement apparent.
b- Aire 6 (aire psycho - motrice) :
En avant de l’aire 4 se trouve un vaste territoire cortical moteur correspondant à l’aire 6, appelée aire pré-motrice. C’est une zone d’intégration motrice qui correspond au point de départ du faisceau fronto-pontin. Elle concerne la commande du mouvement volontaire global (et coordonné). Elle est donc impliquée dans la construction des chaînes musculaires cinétiques assurant le support et la conduite du mouvement volontaire vers son objectif de précision.
c - Aire 8 (aire oculo-motrice) :
En avant de l’aire 6, il existe un petit territoire moteur spécialisé qui est l’aire 8 ou aire oculo-motrice responsable des mouvements synergiques des yeux.
d- Aires 5 et 7
Ce sont des territoires moteurs corticaux associatifs au niveau du lobe pariétal, donnant le faisceau pariéto - pontin. Ces aires sont en relation étroite de proximité avec les aires pariétales de la somesthésie, puis avec l’aire 40 dans la région du pli courbe. Le résultat de cette intégration assure le déroulement normal du mouvement par reconnaissance de l’espace extra - corporel.
Sa pathologie, dans l’hémisphère dominant, entraîne une méconnaissance spatiale du mouvement et une hémi - négligence motrice, du côté opposé à la lésion.
e- Aires 21 et 22.
Ce sont des territoires moteurs corticaux associatifs au niveau du lobe temporal, mais en relation de proximité avec les aires visuelles du lobe occipital. Ces territoires donnent le faisceau temporo-pontin. Ce dernier intervient dans les ataxies à prédominance axiale et, indirectement, sur les mouvements cortico-oculo-céphalogyres (aire 22 : - Bossy p.237).
Le lobe temporal est aussi le lieu des foyers d’épilepsie temporale.
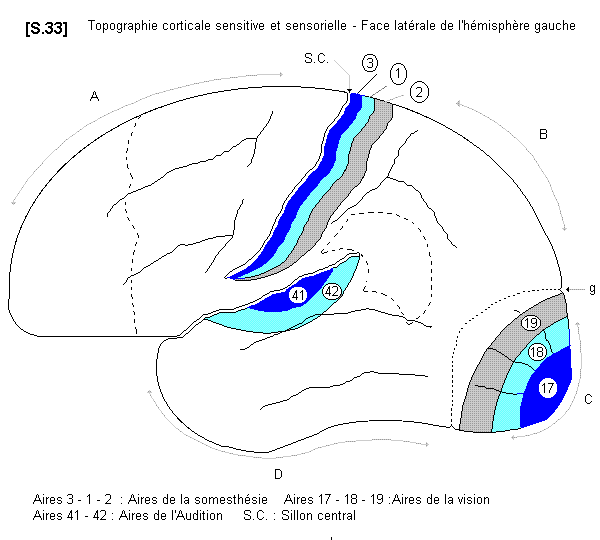
4.4.2.1.2. - fonctions sensitives et sensorielles
....................................................
La topographie sensitive et sensorielle du cortex cérébral a des caractères très particuliers. On distingue en principe trois localisations concentriques pour une même projection sensitive et sensorielle.
a) - Il existe une zone de réception primaire corticale où émerge le signal neuro-physiologique (provenant de la rétine, par exemple).
b) - Autour de la zone primaire, sont disposés des neurones de structure particulière qui correspondent à la prise de conscience de la nature du signal et de ses paramètres. C’est la zone de perception.
c) - Enfin, à la périphérie, il existe une troisième zone de neurones spécialisés qui correspondent à l’analyse du message sensoriel, à son identification et à sa reconnaissance. C’est la zone d’interprétation.
En gros, il existe, en principe, trois centres concentriques qui sont : le centre de réception primaire, le centre de perception consciente et le centre d’interprétation. Les territoires qui sont placés autour du centre de la réception primaire sont appelés zones psychiques ou zones de gnosie. Ce sont des territoires associatifs et intégratifs.
En bref, il existe une diffusion corticale du message sensoriel, qui s’accompagne d’un changement de nature par intellectualisation et enrichissement symbolique. A noter que les déficits sensoriels (ex. cécité) peuvent être partiellement compensés par extension fonctionnelle des zones de gnosie des territoires sensitifs environnants (somesthésie tactile, spatiale, et audition). Il existe donc, au niveau cortical, des suppléances possibles qui, bien que partielles, doivent pouvoir être développées par des procédés éducatifs appropriés. (Exemple : Chez l’aveugle, la perception du monde extérieur peut - être enrichie par le développement très précoce des perceptions non - visuelles : tactiles, auditives, olfactives).
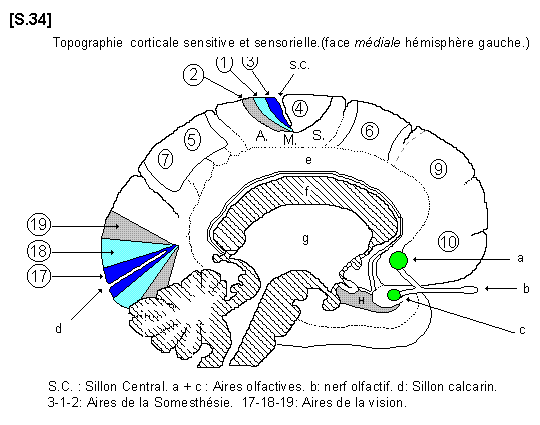
a - Les aires corticales de la vision :
On distingue ainsi dans le lobe occipital les aires sensorielles de la vision. L’aire 17 est le centre de réception primaire, l’aire 18 est l’aire de perception et l’aire 19, la plus périphérique, est l’aire d’interprétation. Ces trois territoires se retrouvent sur la face médiale de l’hémisphère de part et d’autre du sillon calcarin.
b- Les aires corticales de l’audition
Les centres sensoriels de l’audition sont placés au niveau des premières circonvolutions du lobe temporal. On distingue ici seulement deux territoires : l’aire 41 est l’aire de réception primaire et l’aire 42 correspond aux zones de gnosie.
c - Les aires corticales de la somesthésie
La sensibilité générale se projette au niveau du cortex du lobe pariétal, en arrière du sillon central (scissure de Rolando). On distingue là aussi à son niveau, trois territoires successifs : l’aire 3 correspond à la réception primaire et les aires 1 et 2 correspondent aux aires psychiques. Il existe au niveau de ce territoire, sur le gyrus post-central (circonvolution pariétale ascendante) une topographie somatotopique tout à fait comparable à celle de l’homonculus de Penfield. Ce sont aussi des centres associatifs de la reconnaissance des formes et de leur disposition spatiale.
d - Les aires corticales de l’olfaction :
Les aires de l’olfaction sont placées sur la face médiale de l’hémisphère au niveau de la circonvolution limbique et au niveau de la région de l’hippocampe. On distingue deux territoires olfactifs situés dans l’aire 24 et dans l’aire 38, qui correspondent aux deux racines du tractus olfactif.
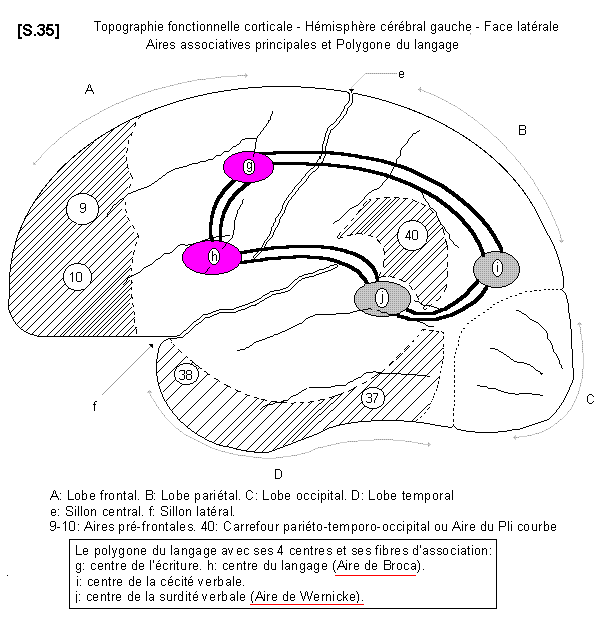
4.4.2.1.3. - Fonctions associatives S.35
Elles couvrent environ 80% du cortex chez l’homme (10% chez le rat), ce qui témoigne de leur importance évolutive et fonctionnelle.
Fonctions générales des aires associatives :
Elles traitent les informations de toutes les aires spécifiques motrices, sensitives et sensorielles. Elles assument ainsi l’intégration de nombreuses informations différentes (par leur nature et leurs paramètres neurophysiologiques). Cette capacité de traitement des signaux aboutit à la construction de programmes de reconnaissance et d’action sur l’environnement. Les conséquences fonctionnelles des déficits pathologiques dans ces territoires corticaux sont des agnosies et des apraxies.
a - Les aires corticales pré - frontales :
Il existe, au niveau du cortex, une zone très particulière, située dans la partie la plus ventrale des lobes frontaux. Ce sont les aires pré - frontales 9 et 10. Leur signification physiologique précise est mal connue. On sait seulement qu’elles représentent le lieu d’élaboration des caractères de la personnalité. Elles concernent aussi les activités intellectuelles de la concentration et de la prévision, ainsi que la régulation de l’humeur (cyclo-thymie : dépression, euphorie). Il s’agit de territoires complexes qui sont en fait des aires associatives, abondamment reliées au lobe limbique, au thalamus et à l’hypothalamus.
Les lésions pré - frontales se caractérisent donc par des modifications caractérielles : perte du dynamisme naturel, perte la concentration, des capacités prévisionnelles, perte de l’initiative et de la décision et perte de l’affectivité, avec conservation des aptitudes intellectuelles fondamentales (raisonnement). En bref, il s’agit d’un état de passivité et de neutralité psycho-affective.
(Etiologies : Traumatismes crânio - cérébraux frontaux, Méningiome frontal opéré).
b - Les aires corticales du polygone du langage
On distingue enfin au niveau du cortex des territoires particuliers sensoriels et moteurs qui sont spécialisés dans les fonctions du langage et de l’écriture (centres du langage et de l’écriture.) On distingue :
* - des centres moteurs. Ce sont :
* le centre de l’écriture : il est placé en avant de la circonvolution frontale ascendante au voisinage des territoires moteurs qui correspondent au membre supérieur et à la main. Sa lésion pathologique provoque l’agraphie.
* le centre du langage : il est situé au pied de la circonvolution frontale ascendante, au voisinage des territoires moteurs du larynx. C’est l’aire de Broca de l’hémisphère gauche, située dans le territoire artériel de la portion initiale de l’artère cérébrale moyenne (ou art. sylvienne) Il s’agit de la composante motrice du langage. Sa lésion provoque l’anarthrie (impossibilité d’émettre des sons organisés). C’est un langage lent, désorganisé, laborieux, en style télégraphique élémentaire, appelé en clinique « locution non fluente » ou « jargon ».
* - des centres de reconnaissance :
* le centre de la surdité verbale : il est situé dans le lobe temporal, en arrière des centres de projection de l’audition. C’est l’aire de Wernicke de l’hémisphère gauche, située dans le territoire artériel de la portion distale de l’artère cérébrale moyenne (ou a. sylvienne). Il s’agit de la composante cognitive du langage, qui concerne le sens du langage. Sa lésion entraîne l’incompréhension des paroles entendues, car elles ne sont pas reconnues. Il s’agit, en fait d’un trouble complexe du langage, associant diverses formes (logorrhée, paraphasie, erreur du mot, manque ou recherche du mot) appelé Aphasie de Wernicke, par lésion dans le lobe temporal postérieur. Mais l’aphasie de Wernicke donne un langage qui parait normal dans sa construction, et qui appelé en clinique « locution fluente ».
Les accidents vasculaires cérébraux dans le territoire de l’artère cérébrale moyenne sont souvent responsables de nécrose ischiémique corticale (ou ramollissent cérébral) au niveau des aires de Broca et de Wernicke.
* le centre de la cécité verbale : il est situé dans une zone intermédiaire entre le lobe pariétal et le lobe occipital. Sa lésion entraîne l’incompréhension des mots écrits ou imprimés, car ils ne sont pas reconnus.
L’ensemble de ces quatre centres constitue le polygone du langage. Il existe entre eux de nombreuses fibres d’association (dont le faisceau arqué, situé dans le territoire du pli courbe) qui relie les aires de Wernicke et de Broca). En effet, on entend et on comprend ses propres paroles, de même que l’on voit et que l’on comprend sa propre écriture. Il faut noter que ce polygone du langage, bien que bilatéral, n’est fonctionnel qu’à gauche chez les droitiers (et à droite chez les gauchers). Pour expliquer cette fonction unilatérale, on pense que les phénomènes moteurs et psychiques du langage, dont le synchronisme est très délicat, nécessitent la prédominance d’un hémisphère sur l’autre. C’est ainsi que s’expliqueraient certaines formes de bégaiement par non - prédominance d’un hémisphère sur l’autre.
Ces constatations posent le problème plus général de la spécialisation fonctionnelle des aires associatives de chacun des hémisphères.
Les aires associatives de l’hémisphère dominant (le gauche chez les droitiers) paraissent plus aptes aux fonctions procédurales, c’est-à-dire au traitement séquentiel et analytique des informations et, par conséquence, plus aptes aux raisonnements déductifs et logiques (les mathématiques par exemple).
Au contraire, les mêmes territoires dans l’autre hémisphère, paraissent plus aptes aux fonctions déclaratives, c’est à dire au traitement global et intuitif des informations, considérées comme un tout. Ce serait le lieu de la reconnaissance analogique des images sensorielles - visuelles - sonores - olfactives - gustatives (notion de l’espace environnant, des formes, des couleurs, de la musique, et de leurs nuances).
c - Le carrefour temporo-pariéto-occipital :
Il s’agit d’une zone associative très importante située à cheval sur trois lobes : le lobe pariétal, le lobe temporal et le lobe occipital et qui correspond à l’aire 40. C’est la région du pli courbe. Cette aire est principalement le territoire de la représentation spatiale extérieure et du schéma corporel. Ses troubles pathologiques se caractérisent par des déficits fonctionnels complexes, qui sont :
a) - des apraxies :
perturbations de l’exécution motrice volontaire et coordonnée.
b) - des agnosies :
troubles de l’identification et de la reconnaissance.
En bref, cette région est un grand centre de synthèse d’informations différentes.
d - Les aires associatives du lobe temporal
En plus des aires auditives et des aires motrices extra-pyramidales (origine du faisceau temporo-pontique), le cortex temporal contient de vastes territoires associatifs qui correspondent aux circonvolutions T3 et T4 ( aires 36, 37, 38 ), et concernent la mémorisations d’informations, spécialement visuelles.
A noter que T5 appartient fonctionnellement au système limbique.
4.4.2.2. - Systématisation du paléencéphale
Noyaux gris centraux - Centres et voies motrices extra - pyramidales S.36
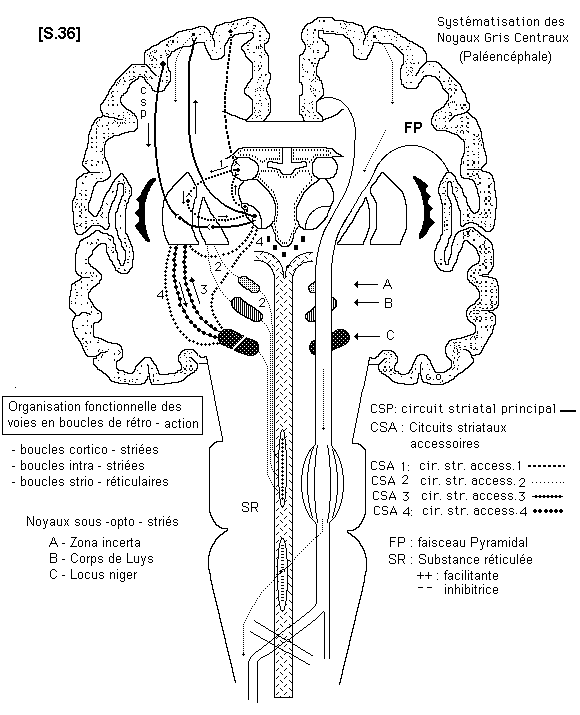
Les noyaux gris centraux sont constitués par le corps strié et la couche optique ou thalamus. Ces structures ont été étudiées au chapitre de la morphologie. Ils jouent un rôle important dans la fonction motrice et dans le contrôle de son déroulement. En effet, toute activité motrice n’est pas volontaire. Il existe à l’intérieur du mouvement normal une grande participation d’activités motrices automatiques qui résultent très principalement des fonctions des noyaux gris centraux, situés dans le paléencéphale. Leurs voies neurologiques particulières constituent les voies motrices extra - pyramidales dont les faisceaux de sortie atteignent plus bas, tous les segments de la moelle épinière.
Au niveau des noyaux gris centraux, ces voies constituent les circuits striataux.
°Les circuits striataux
Les différentes connexions des noyaux gris centraux entre eux et avec le cortex, décrivent plusieurs systèmes de boucles de régulation de l’activité motrice, dont la participation est indispensable à l’exécution normale du mouvement. Ces connexions sont assurées par des voies et des synapses qui mettent en jeu des neuro-médiateurs et des récepteurs spécifiques.
On distingue actuellement 5 boucles de régulation :
- le circuit striatal principal (CSP) : il est cortico-strio-thalamo-cortical.
- les quatre circuits striataux accessoires (CSA) :
* 1er CSA : néo-striatum, pallidum, noyau latéro-ventral intermédiaire du thalamus et retour au cortex.
* 2è CSA : boucle pallido-sous-thalamo-striée : elle relie le pallidium au corps de Luys et à la zona incerta.
* 3è CSA : c’est la boucle strio-nigrique qui est impliquée dans la physio-pathologie de la maladie de Parkinson.
* 4è CSA : boucle strio-nigro-thalamique et retour au néo-striatum et retour au néo-striatum.
(A noter, l’organisation générale en boucles de contrôle ou de rétro - action).
° les systèmes d’entrée dans les circuits striataux :
Le cortex du cerveau projette des fibres sur le paléencéphale (sur le néo-striatum puis, au delà, sur le pallidum). Ce sont les voies cortico -striées.
° les systèmes intra - striataux :
C’est le circuit striatal accessoire 1. Il relie le néo- striatum (Noyau caudé/Putamen) au paléo - striatum (Pallidum/noyaux latéro - ventraux du thalamus).
° les systèmes de sortie des circuits striataux :
Les corps striés sont en relation avec les noyaux sous-jacents : noyaux sous opto -striés et, au delà, avec la substance réticulée. Ce sont les voies strio - réticulaires.
Physio - pathologie du paléencéphale (Pathologie motrice extra - pyramidale)
Applications cliniques :
La pathologie extra pyramidale comporte les signes cliniques suivants :
- troubles du tonus musculaire : l’hypertonie est une rigidité de type plastique. Elle est, en principe, diffuse. Cependant une hypertonie localisée entraîne une dystonie, responsable de mouvements anormaux, incontrôlables. Ils se voient dans l’Athétose, la Chorée, et l’Hémiballisme. Ce dernier correspond à une lésion sélective du Corps de Luys.
- tremblements rythmiques de repos qui disparaissent au mouvement.
- akinésie par difficulté d’exécution du mouvement volontaire, aboutissant à la rareté du geste et de l’expression (Maladie de Parkinson).
En bref, les lésions des centres et des voies du paléencéphale se traduisent par des troubles du mouvement et des troubles du tonus musculaire. On distingue ainsi deux types de syndromes extra -pyramidaux :
– Un syndrome hypokinétique - hypertonique (rigidité musculaire) :
Ex : Maladie de Parkinson (boucle strio - nigrique)
– Un syndrome hyperkinétique - hypotonique : Ex : Athétose (lésion du néo - striatum).
Pr Gérard Outrequin - Dr Bertrand Boutillier - Toute reproduction interdite
Licence d'utilisation - Mentions légales - Politique de confidentialité - Découvrez MedShake !